
Eurypterid: Difference between revisions
m Undid revision 819976924 by Ichthyovenator (talk) Actually it didnt |
Rescuing 2 sources and tagging 1 as dead. #IABot (v1.6.2) (Balon Greyjoy) |
||
| Line 111: | Line 111: | ||
=== Locomotion === |
=== Locomotion === |
||
[[File:Subaqueous flight.png|thumb|Illustration of subaqueous flight in ''[[Eurypterus]]'' in which the shape of the paddles and their motion through water is enough to generate [[Lift (force)|lift]].]] |
[[File:Subaqueous flight.png|thumb|Illustration of subaqueous flight in ''[[Eurypterus]]'' in which the shape of the paddles and their motion through water is enough to generate [[Lift (force)|lift]].]] |
||
The locomotion of the eurypterids varied greatly from genus to genus. The legs of many genera were far too small to do much more than allow them to crawl across the sea bottom.<ref name=":0">{{Cite web|url= |
The locomotion of the eurypterids varied greatly from genus to genus. The legs of many genera were far too small to do much more than allow them to crawl across the sea bottom.<ref name=":0">{{Cite web|url=http://www.palaeos.com/Invertebrates/Arthropods/Chelicerata/Eurypterida/Eurypterida.html|title=Palaeos Metazoa: Arthropoda: Chelicerata: Eurypterida: Eurypterida|date=2007-08-13|access-date=2017-12-17|deadurl=yes|archiveurl=https://web.archive.org/web/20070813065212/http://www.palaeos.com/Invertebrates/Arthropods/Chelicerata/Eurypterida/Eurypterida.html|archivedate=2007-08-13|df=}}</ref> This is especially the case in the suborder [[Eurypterina]]. Eurypterines and [[Stylonurina|Stylonurines]] (the other suborder) are separated by the morphology of the posteriormost prosomal appendage. In the Stylonurina, this appendage takes the form of a long and slender walking leg whilst in the Eurypterina, the leg is most usually modified and broadened into a swimming paddle.<ref name="tetlie20072">{{cite journal | first = O. Erik | last = Tetlie | name-list-format = vanc | year=2007|title=Distribution and dispersal history of Eurypterida (Chelicerata)|url=http://fossilinsects.net/pdfs/tetlie_2007_PX3_DistribDispersalEurypterida.pdf|deadurl=yes|format=[[Portable Document Format|PDF]]|journal=[[Palaeogeography, Palaeoclimatology, Palaeoecology]]|volume=252|issue=3–4|pages=557–574|doi=10.1016/j.palaeo.2007.05.011|archiveurl=https://web.archive.org/web/20110718202232/http://fossilinsects.net/pdfs/tetlie_2007_PX3_DistribDispersalEurypterida.pdf|archivedate=2011-07-18|df=}}</ref> |
||
Unlike the Eurypterines, a number of Stylonurines had elongated and powerful legs that would clearly have allowed them to walk even on land (not unlike modern [[Crab|crabs]]).<ref name=":0" /> Studies of what are believed to be [[fossil trackway]]s indicate that eurypterids used in-phase, hexapodous (six-legged) and octopodous (eight-legged) gaits.<ref>{{cite journal | vauthors = Whyte MA | title = Palaeoecology: a gigantic fossil arthropod trackway | journal = [[Nature (journal)|Nature]]| | volume = 438 | issue = 7068 | pages = 576 | date = December 2005 | pmid = 16319874 | doi = 10.1038/438576a }}</ref> Some species may have been amphibious, emerging onto land for at least part of their life cycle; they may have been capable of breathing both in water and in air. |
Unlike the Eurypterines, a number of Stylonurines had elongated and powerful legs that would clearly have allowed them to walk even on land (not unlike modern [[Crab|crabs]]).<ref name=":0" /> Studies of what are believed to be [[fossil trackway]]s indicate that eurypterids used in-phase, hexapodous (six-legged) and octopodous (eight-legged) gaits.<ref>{{cite journal | vauthors = Whyte MA | title = Palaeoecology: a gigantic fossil arthropod trackway | journal = [[Nature (journal)|Nature]]| | volume = 438 | issue = 7068 | pages = 576 | date = December 2005 | pmid = 16319874 | doi = 10.1038/438576a }}</ref> Some species may have been amphibious, emerging onto land for at least part of their life cycle; they may have been capable of breathing both in water and in air. |
||
Eurypterines were more clearly aquatic, with their sixth appendage developed into a broad swimming paddle. The swimming paddles could only be moved in near-horizontal planes (not upwards or downwards).<ref name="autec">{{cite journal| |
Eurypterines were more clearly aquatic, with their sixth appendage developed into a broad swimming paddle. The swimming paddles could only be moved in near-horizontal planes (not upwards or downwards).<ref name="autec">{{cite journal|vauthors=Selden PA|year=1999|title=Autecology of Silurian Eurypterids|url=http://homepage.mac.com/paulselden/Sites/Website/Autecology.pdf|journal=Special Papers in Palaeontology|publisher=Palaeontological Association|volume=32|pages=39–54|issn=0038-6804|accessdate=May 23, 2011}}{{dead link|date=January 2018 |bot=InternetArchiveBot |fix-attempted=yes }}</ref> As such, swimming Eurypterids are generally agreed to have utilized a rowing type of propulsion similar to that of [[Crab|crabs]] and [[Water beetle|water beetles]].<ref name="merck">{{cite web|url=http://www.geol.umd.edu/~jmerck/bsci392/lecture10/lecture10.html|title=The biomechanics of swimming | first = John W. | last = Merck | name-list-format = vanc |publisher=Department of Geology, University of Maryland|accessdate=May 23, 2011}}</ref> Larger individuals may have been capable of underwater flying (or [[Aquatic locomotion|subaqueous flight]]) in which the motion and shape of the paddles are enough to generate [[Lift (force)|lift]], similar to the swimming of [[Sea turtle|sea turtles]] and [[Sea lion|sea lions]]. It has a relatively slower acceleration rate than the rowing type, especially since adults have proportionally smaller paddles than juveniles. But since the larger sizes of adults mean a higher [[drag coefficient]], using this type of propulsion is more energy-efficient.<ref name="autec" /><ref name="merck2">{{cite web|url=http://www.geol.umd.edu/~jmerck/bsci392/lecture25/lecture25.html|title=Eurypterids, arachnids, and the arthropod invasion of the land| first = John W. | last = Merck | name-list-format = vanc | publisher=Department of Geology, University of Maryland|accessdate=May 23, 2011}}</ref> |
||
=== Sexual dimorphism and reproduction === |
=== Sexual dimorphism and reproduction === |
||
| Line 198: | Line 198: | ||
* {{cite book | first1 = John M. | last1 = Clarke | first2 = R. | last2 = Rudolf | name-list-format = vanc |year=1912 |title=The Eurypterida of New York |location=Albany, New York |publisher=[[New York State Education Department]]}} |
* {{cite book | first1 = John M. | last1 = Clarke | first2 = R. | last2 = Rudolf | name-list-format = vanc |year=1912 |title=The Eurypterida of New York |location=Albany, New York |publisher=[[New York State Education Department]]}} |
||
* {{cite journal | first1 = Neal S. | last1 = Gupta | first2 = O. Erik | last2 = Tetlie | first3 = Derek E. G. | last3 = Briggs | first4 = Richard D. | last4 = Pancost | name-list-format = vanc |year=2007 |title=The fossilization of eurypterids: a result of molecular transformation |journal=[[Palaios]] |volume=22 |issue=4 |pages=439–447 |url=http://paleo.ku.edu/palaios/22/4/gupta.pdf |format=[[Portable Document Format|PDF]] |doi=10.2110/palo.2006.p06-057r}} |
* {{cite journal | first1 = Neal S. | last1 = Gupta | first2 = O. Erik | last2 = Tetlie | first3 = Derek E. G. | last3 = Briggs | first4 = Richard D. | last4 = Pancost | name-list-format = vanc |year=2007 |title=The fossilization of eurypterids: a result of molecular transformation |journal=[[Palaios]] |volume=22 |issue=4 |pages=439–447 |url=http://paleo.ku.edu/palaios/22/4/gupta.pdf |format=[[Portable Document Format|PDF]] |doi=10.2110/palo.2006.p06-057r}} |
||
* {{cite journal | |
* {{cite journal |first1=Phillip L. |last1=Manning |first2=Jason A. |last2=Dunlop |name-list-format=vanc |year=1995 |title=The respiratory organs of eurypterids |journal=[[Palaeontology (journal)|Palaeontology]] |volume=38 |issue=2 |pages=287–297 |url=http://palaeontology.palass-pubs.org/pdf/Vol%2038/Pages%20287-297.pdf |format=[[Portable Document Format|PDF]] |deadurl=yes |archiveurl=https://web.archive.org/web/20120309062653/http://palaeontology.palass-pubs.org/pdf/Vol%2038/Pages%20287-297.pdf |archivedate=2012-03-09 |df= }} |
||
* {{cite journal | first = O. Erik | last = Tetlie | name-list-format = vanc |year=2007 |title=Distribution and dispersal history of Eurypterida (Chelicerata) |journal=[[Palaeogeography, Palaeoclimatology, Palaeoecology]] |volume=252 |issue=3–4 |pages=557–574 |url=http://fossilinsects.net/pdfs/tetlie_2007_PX3_DistribDispersalEurypterida.pdf |format=[[Portable Document Format|PDF]] |doi=10.1016/j.palaeo.2007.05.011 |deadurl=yes |archiveurl=https://web.archive.org/web/20110718202232/http://fossilinsects.net/pdfs/tetlie_2007_PX3_DistribDispersalEurypterida.pdf |archivedate=2011-07-18 |df= }} |
* {{cite journal | first = O. Erik | last = Tetlie | name-list-format = vanc |year=2007 |title=Distribution and dispersal history of Eurypterida (Chelicerata) |journal=[[Palaeogeography, Palaeoclimatology, Palaeoecology]] |volume=252 |issue=3–4 |pages=557–574 |url=http://fossilinsects.net/pdfs/tetlie_2007_PX3_DistribDispersalEurypterida.pdf |format=[[Portable Document Format|PDF]] |doi=10.1016/j.palaeo.2007.05.011 |deadurl=yes |archiveurl=https://web.archive.org/web/20110718202232/http://fossilinsects.net/pdfs/tetlie_2007_PX3_DistribDispersalEurypterida.pdf |archivedate=2011-07-18 |df= }} |
||
* {{cite journal | first1 = O. Erik | last1 = Tetlie | first2 = Michael B. | last2 = Cuggy | name-list-format = vanc |year=2007 |title=Phylogeny of the basal swimming eurypterids (Chelicerata; Eurypterida; Eurypterina) |journal=[[Journal of Systematic Palaeontology]] |volume=5 |pages=345–356 |doi=10.1017/S1477201907002131 |issue=3}} |
* {{cite journal | first1 = O. Erik | last1 = Tetlie | first2 = Michael B. | last2 = Cuggy | name-list-format = vanc |year=2007 |title=Phylogeny of the basal swimming eurypterids (Chelicerata; Eurypterida; Eurypterina) |journal=[[Journal of Systematic Palaeontology]] |volume=5 |pages=345–356 |doi=10.1017/S1477201907002131 |issue=3}} |
||
Revision as of 19:40, 22 January 2018
| Eurypterida Temporal range: Darriwilian-Late Permian,
| |
|---|---|

| |
| Eurypterid from Ernst Haeckel's Kunstformen der Natur (1904) | |
| Scientific classification | |
| Kingdom: | |
| Phylum: | |
| Subphylum: | |
| Class: | |
| Order: | † Eurypterida Burmeister, 1843
|
| Suborders | |
|
†Stylonurina Diener, 1924 | |
| Synonyms | |
| |
Eurypterids, often informally called sea scorpions, are an extinct group of arthropods related to arachnids that include the largest known arthropods to have ever lived. They are members of the extinct order Eurypterida (Chelicerata); which is the most diverse Paleozoic chelicerate order in terms of species.[1] The name Eurypterida comes from the Greek words eury- (meaning "broad" or "wide") and pteron (meaning "wing").[2] This name was chosen due to the pair of wide swimming appendages on the first fossil eurypterids discovered. The largest, such as Jaekelopterus, reached 2.5 metres (8 ft 2 in) in length, but most species were less than 20 centimetres (8 in). They were formidable predators that thrived in warm shallow water, in both seas and lakes,[3] from the mid Ordovician to late Permian (467.3 to 252 million years ago).
Although called "sea scorpions", only the earliest ones were marine (later ones lived in brackish or freshwater), and they were not true scorpions. According to theory, the move from the sea to fresh water had probably occurred by the Pennsylvanian subperiod. Some studies suggest that a dual respiratory system was present, which would allow short periods of time in terrestrial environments.[4] Eurypterids are believed to have undergone ecdysis, making their significance in ecosystems difficult to assess, because it can be difficult to tell a fossil moult from a true fossil carcass.[5] They became extinct during the Permian–Triassic extinction event or sometime before the event 251.902 million years ago. Their fossils have a near global distribution.
About two dozen families of eurypterids are known. Perhaps the best-known genus of eurypterid is Eurypterus, of which around 16 fossil species are known. The genus Eurypterus was described in 1825 by James Ellsworth De Kay, a zoologist. He recognized the arthropod nature of the first-ever described eurypterid specimen, found by Dr. S. L. Mitchill. In 1984, that species, Eurypterus remipes was named the state fossil of New York.
Pentecopterus decorahensis, which lived as early as 467.3 million years ago, is the oldest described eurypterid; with an estimated length of 1.83 metres (6 ft 0 in),[6][7] it has been described as "the first real big predator".[8][9]
Morphology

Eurypterids have been formally described as follows:
Small to very large merostomes with elongate lanceolate, rarely trilobed body; prosoma [head] of moderate size; opisthosoma [body] with 12 moveable segments and styliform to spatulate telson [tail], with division commonly into 7-segmented preabdomen and 5-segmented postabdomen; prosomal [head] appendages 6, comprising 3-jointed chelicerae, walking legs, the last pair commonly transformed into swimming legs. Mouth central, bordered posteriorly by endostoma and metastoma. Operculum with median genital appendage, abdominal appendages plate-shaped with nonlaminate gills. Ordovician-Permian.[10]
The typical eurypterid had a large, flat, semicircular carapace, followed by a jointed section, and finally a tapering, flexible tail, most ending with a long spine at the end (Pterygotus, though, had a large flat tail, possibly with a smaller spine). As with the xiphosurans, the eurypterid body can be divided into two primary sections. The frontal part (or "head" part) is known as the prosoma. The legs, eyes and mouth are all located on the prosoma and it serves as both a head and a thorax. The upper side of the prosoma holds the compound eyes and between them a pair of small ocelli (simple eye-like sensory organs). The mouth is on the underside and is surrounded by six pairs of legs. Gnathobases, or "toothplates" connect the legs to the body and are lined with teeth.[11]

The modified first pair of legs are known as chelicerae (differently developed, but the same organs as spider fangs). In the pterygotids, these and their claws were enlarged. Following the first pair, the four subsequent pairs of legs are walking legs. These legs are cylindrical, spiny in some species, and tend to get larger the further back they are. The sixth pair of appendages are differently developed in the two different eurypterid suborders. In the Stylonurina, they serve as an additional pair of walking legs. In the Eurypterina, they are flattened and paddle-like to be used for swimming. The endostoma, a U-shaped and small plate, borders the mouth and is covered by the metastoma, a larger plate and part of the abdomen.[11]
Behind the head, eurypterids had twelve body segments. This posterior section, longer than the frontal section, is called the opisthosoma. Due to its length it is often divided further into the pre-abdomen (consisting of seven segments) and a more narrow post-abdomen (consisting of five segments). Some researchers divide it into the mesosoma (first six segments) and metasoma (last six segments) instead. Each segment was formed from a dorsal plate (tergite) and a ventral plate (sternite). The last segment, or "tail", possesses a telson, most often a blade-like spine (through other shapes exist). It may have been used to inject venom in some species, though there is no evidence that eurypterids were venomous.[11]
The largest eurypterid and the largest known arthropod ever to have lived is Jaekelopterus rhenaniae supported by the finding of a 46 cm (18 in) long claw in 2007, indicating a body length of 2.5 m (8 ft 2 in).[12] Other large eurypterids include Pterygotus, with the largest species reaching lengths of about 1.6 m[12], Pentecopterus decorahensis with an estimated length of 1.83 m[13] and Hibbertopterus scouleri with a length potentially just short of 2 m.[14] Many smaller eurypterids were still likely formidable predators, with sizes around 100 cm being common in most eurypterid clades.[4]
Evolutionary history
The earliest known eurypterids - Brachyopterus and Pentecopterus - are from the Middle Ordovician, and the last recorded members of the group - Campylocephalus - are from the Late Permian, giving a total range of over 200 million years. They were most diverse between the Middle Silurian and Early Devonian, peaking in diversity during the Late Silurian.[4]
Origins

The origin of the eurypterids is obscure. Though "primitive eurypterids" have on occasion been described from deposits of Cambrian or even Pre-Cambrian age,[15] these fossils are not recognized as eurypterids (and sometimes not even as related forms) today. Some animals previously seen as "primitive eurypterids", such as the genus Strabops from the Cambrian of Missouri[15], are now classified as aglaspidids. The aglaspidids, once seen as primitive chelicerates are now seen as a group more closely related to trilobites.[16]
A predatory arthropod known as Protichnites,[17] whose traces are found in Cambrian strata dating from 510 million years ago, is a possible stem group eurypterid, and is among the first evidence of animals on land.[18]
The appearance of several morphologically different clades of eurypterids during the Darriwilian stage of the Ordovician indicate that the eurypterids experienced an explosive radiation during the Early Ordovician or that they first appeared during the Cambrian.[19]
Ordovician

Until 1882 no eurypterids were known from before the Silurian, though discoveries throughout the twentieth century and modern times have expanded the knowledge of early eurypterids from the Ordovician period.[20] The earliest eurypterids known today date from the Middle Ordovician.[4] There are reports of fossil eurypterids in deposits of Tremadocian (Early Ordovician) age in Morocco, but these have yet to be thoroughly studied.[21]
Both major eurypterid groups, the Stylonurina and Eurypterina, had already been established 460 million years ago. The presence of members of both groups, notably Stylonurine Brachyopterus[4] and Eurypterine Pentecopterus[19], indicate that primitive stem-eurypterids would have preceeded them, though these are so far unknown in the fossil record. Furthermore, the phylogenetic positions of the few Ordovician eurypterids known indicate that they must have been very diverse during this early period of their evolution, despite being rare in the fossil record.[22]
Indeed, the fossil record of Ordovician eurypterids is very poor. The majority of eurypterids once reportedly known from the Ordovician have since proven to be misidentifications or pseudofossils. Today only 11[19][23] species can confidently be identified as representing Ordovician eurypterids. These taxa fall into two distinct ecological categories; large and active predators of Laurentia and demersal and basal animals from Avalonia and Gondwana.[19] The Megalograptoidea of Laurentia likely represent the first truly successful eurypterid group, experiencing a radiation during the Late Ordovician.[4]
Some genera of eurypterids appearing during the Ordovician period include[19]:
Silurian
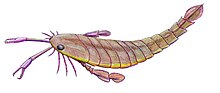
Of the 150-160 species of eurypterids known in 1916 more than half were from the Silurian, with a third from the Late Silurian alone. As such, it has been concluded that the group peaked in number and diversity during the Silurian period.[15] Approximately 100 additional eurypterid species have been described since 1916[23], though there is still a distinct absolute peak in eurypterid diversity seen during the Late Silurian.[4]
A vast majority of eurypterid groups are first recorded from the Silurian, among them the Stylonuroidea, Kokomopteroidea, Mycteropoidea, Pterygotioidea, Eurypteroidea and Waeringopteroidea[23], though at least some were likely already present during the Late Ordovician.[19] The Stylonurina remained rare, as they had been during the Ordovician, but the Eurypterina saw a rapid rise in diversity and number.[4]
The Silurian Eurypterus was the most successful eurypterid by far, dominating many marine eurypterid faunas and accounting for more than 90 % of all known eurypterid specimens.[4] The Pterygotioidea, large predatory forms, achieved a cosmopolitan distribution, but would not survive for long, going extinct during the Early Devonian.[4]
Some genera of eurypterids appearing during the Silurian period include[23]:
Devonian

The eurypterids were one of many groups affected by the Late Devonian extinction. A major decline in diversity had began during the Early Devonian and they were rare in marine environments by the Late Devonian. Elevated extinction rates have been observed during the Frasnian (extinction of four families) and Famennian (extinction of five families) stages. The families surviving into the Carboniferous were all freshwater groups.[24] Though the diversity and number of the Eurypterina waned, a slow extinction possibly tied to the emergence of jawed vertebrates, the Stylonurina were relatively unaffected, adapting new strategies (such as sweep-feeding) to avoid competition.[25]
The radiation of the Stylonurine Hibbertopteridae in the Late Devonian and Carboniferous was the last major radiation of eurypterids before their extinction in the Permian.[26]
Some genera of eurypterids appearing during the Devonian period include[23]:
Carboniferous and Permian

By the Carboniferous, only three families of eurypterids remained. The Stylonurines survived in the Hibbertopteridae and Mycteroptidae[26], whilst the Eurypterina was represented by a single genus, Adelophthalmus.[4] Both Carboniferous Stylonurine families and Adelophthalmus would last into the Permian.[23]
Adelophthalmus, an able swimmer, was already widespread during the Devonian, with fossils having been found in both Siberia and Australia. With the amalgamation of Pangaea during the Carboniferous and Permian, the genus gained an almost cosmopolitan distribution.[4]
No eurypterids are known from fossil beds higher than the Permian, indicating that the last eurypterids died either during the period or in the catastrophic extinction event at its end. The Permian-Triassic extinction event was the most devastating mass extinction event recorded and also rendered many other Paleozoic groups, such as the trilobites, extinct.[27] The last known surviving eurypterid was the Late Permian Campylocephalus permianus, recovered from deposits in Russia about 250 million years old.[4]
Some genera of eurypterids appearing during the Carboniferous and Permian periods include[23]:
Biology
Locomotion

The locomotion of the eurypterids varied greatly from genus to genus. The legs of many genera were far too small to do much more than allow them to crawl across the sea bottom.[11] This is especially the case in the suborder Eurypterina. Eurypterines and Stylonurines (the other suborder) are separated by the morphology of the posteriormost prosomal appendage. In the Stylonurina, this appendage takes the form of a long and slender walking leg whilst in the Eurypterina, the leg is most usually modified and broadened into a swimming paddle.[28]
Unlike the Eurypterines, a number of Stylonurines had elongated and powerful legs that would clearly have allowed them to walk even on land (not unlike modern crabs).[11] Studies of what are believed to be fossil trackways indicate that eurypterids used in-phase, hexapodous (six-legged) and octopodous (eight-legged) gaits.[29] Some species may have been amphibious, emerging onto land for at least part of their life cycle; they may have been capable of breathing both in water and in air.
Eurypterines were more clearly aquatic, with their sixth appendage developed into a broad swimming paddle. The swimming paddles could only be moved in near-horizontal planes (not upwards or downwards).[30] As such, swimming Eurypterids are generally agreed to have utilized a rowing type of propulsion similar to that of crabs and water beetles.[31] Larger individuals may have been capable of underwater flying (or subaqueous flight) in which the motion and shape of the paddles are enough to generate lift, similar to the swimming of sea turtles and sea lions. It has a relatively slower acceleration rate than the rowing type, especially since adults have proportionally smaller paddles than juveniles. But since the larger sizes of adults mean a higher drag coefficient, using this type of propulsion is more energy-efficient.[30][32]
Sexual dimorphism and reproduction
As in many other entirely extinct groups, understanding and researching the reproduction and sexual dimorphism of eurypterids is difficult, as they are only known from fossilized shells and carapaces.[11] Sometimes two sexes of the same species have been interpreted as two different species, as was the case with two species of Drepanopterus (D. bembycoides and D. lobatus).[33] In other cases, there might not be enough apparent differences to differentiate different sexes based on morphology alone.[11]
The eurypterid prosoma is made up of the first six exoskeleton segments fused together. The seventh segment is what is called the metastoma and the eighth plate-like segment (called the operculum) contains the genital aperature. In its center, as in modern Xiphosurans, is a genital appendage. This appendage is an elongated rod and exists in two distinct morphs, both with internal ducts.[11] Which morph belongs to which gender is controversial. "Type A" genital appendages, equipped with clasping organs, have generally been interpreted as male organs whilst the other morph, "type B", has been interpreted as a female ovipositor (used to deposit eggs) that fits over the claspers of the male.[11] It is also possible that reproduction was done through a spermatophore transfer on the substrate and that the "Type A" appendage would be the female morph, used to collect and store the spermatophore in her spermathecae.[11]
History of discovery

The earliest discovered eurypterid fossil was unearthed in the fossil-rich Siluric rocks of New York. Dr S. L. Mitchill errouniously identified the fossil as an example of the fish Silurus in 1818, likely due to the strange, catfish-like appearance of the carapace. Seven years later, in 1825, James E. DeKay recognized the fossil as clearly belonging to an arthropod. He identified it as a crustacean of the order Branchiopoda and concluded that the genus Triops was likely closely related to it. DeKay further suggested that the fossil, which he named Eurypterus remipes, might be a missing link between the trilobites and the branchiopods.[34]
Early descriptions of other eurypterids from New York preceded discoveries in Europe by a considerable amount of time and many european species later recognized as distinct, such as Eurypterus tetragonophthalmus from the Baltic, were first seen as examples of E. remipes.[34]
Nieszkowski's De Euryptero Remipede (1858) featured an extensive description of E. fischeri (now seen as synonymous with E. tetragonophthalmus), which along with the monograph On the Genus Pterygotus by Huxley and Salter and an exhaustive description of the various eurypterids of New York in volume 3 of the Palaeontology of New York (1859) by James Hall contributed massively to the understanding of eurypterid diversity and biology. These publications were the first to fully describe the whole anatomy of eurypterids, recognizing the full number of prosomal appendages and the number of pre-abdominal and post-abdominal segments. Both Nieszkowski and Hall recognized that the eurypterids were closely related to modern horseshoe crabs.[34]
In the work Anatomy and Relations of the Eurypterida (1893), Laurie added considerably to the knowledge and discussion of eurypterid anatomy and relations, focusing on how the eurypterids related to each other and to trilobites, crustaceans, scorpions, other arachnids and horseshoe crabs. The description of Eurypterus fischeri by Holm (1896) was so elaborate that the species became one of the most completely known of all extinct animals, so much so that the knowledge of E. fischeri was comparable with the knowledge of its modern relatives (such as Limulus). The description also helped solidify the close relationship between the eurypterids and the horseshoe crabs by showcasing numerous homologies between the two groups.[34]
Classification
Eurypterids have traditionally been regarded as close relatives of horseshoe crabs (Xiphosura), together forming a group called Merostomata.[35] Subsequent studies placed eurypterids closer to the arachnids in a group called Metastomata.[36]
There has also been a prevailing idea that eurypterids are closely related to scorpions, which they resemble.[37] This hypothesis is reflected in the common name "sea scorpion". More recently it has been recognised that a little-known, extinct group called Chasmataspida also shares features with Eurypterida,[38] and the two groups were sometimes confused with one another.
A recent summary of the relationships between arachnids and their relatives recognised Eurypterida, Xiphosura and Arachnida as three major groups, but was not able to resolve the phylogenetic relationship of any shared details between them.[39] Another suggested the eurypterids were sister group to the chasmataspids, with these two groups in turn sister group to the horseshoe crabs.[35]
Phylogeny

The cladogram presented here is simplified from a study by Tetlie.[40] The most important phylogenetic breakdown is based on the two major innovations that characterise the evolution of the eurypterids. The most important was the transformation of the posteriormost prosomal appendage into a swimming paddle (as found in the clade Eurypterina). The second innovation was the enlargement of the chelicerae, (as found in the family Pterygotidae), allowing these appendages to be used for active prey capture.
Seventy-five percent of eurypterid species are eurypterines; this represents 99% of specimens.[1] The superfamily Pterygotioidea is the most species-rich clade, with 56 species, followed by the Adelophthalmoidea with 43 species; as sister taxa, they comprise the most derived eurypterids. Pterygotioidea includes the pterygotids, which are the only eurypterids to have a cosmopolitan distribution.[40] This clade is one of the best supported within the eurypterids.
It has been suggested that the development of dermal armour in certain groups of jawless vertebrates (such as the Heterostraci and the Osteostraci) is in response to predation pressure by increasingly sophisticated eurypterid predators[41] (specifically the pterygotids) although this has yet to be verified by detailed analysis.[42] An increase in fish diversity is tied to a decline in eurypterid diversity in the Lower Devonian,[43] although it is not thought that this represents competitive replacement; in fact, this is rare in the fossil record.[44]
| Eurypterida | |
See also
- List of eurypterids
- Pentecopterus—the earliest known eurypterid.
- Hibbertopterus—the last known surviving eurypterid.
- Nepidae—an unrelated insect, commonly known as a "water scorpion".
References
- ^ a b Dunlop JA, Penney D, Tetlie OE, Anderson LT (2008). "How many species of fossil arachnids are there?". Journal of Arachnology. 36 (2): 267–272. doi:10.1636/CH07-89.1. ISSN 0161-8202.
{{cite journal}}: Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - ^ Webster's New Universal Unabridged Dictionary. 2nd ed. 1979.
- ^ Hannibal, Joseph T.; Lucas, Spencer G.; Lerner, Allan J.; Chaney, Dan S. (2005). Lucas, S. G.; Zeigler, K. E.; Spielmann, J. A. (eds.). "An eurypterid (Adelophthalmus sp.) from a plant-rich lacustrine facies of Upper Pennsylvanian strata in El Cobre Canyon, New Mexico" (PDF). New Mexico Museum of Natural History and Science Bulletin. The Permian of Central New Mexico. 31: 34–38.
{{cite journal}}: Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - ^ a b c d e f g h i j k l m "Distribution and dispersal history of Eurypterida (Chelicerata)". Palaeogeography, Palaeoclimatology, Palaeoecology. 252 (3–4): 557–574. 2007-09-03. doi:10.1016/j.palaeo.2007.05.011. ISSN 0031-0182.
- ^ Brandt, Danita; McCoy, Victoria (2009). "Scorpion taphonomy: criteria for distinguishing fossil scorpion molts and carcasses". Journal of Arachnology. 37 (3): 312. doi:10.1636/sh09-07.1.
{{cite journal}}: Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - ^ Shelton, Jim (August 31, 2015). "Meet Pentecopterus, a new predator from the prehistoric seas". Yale University. Retrieved September 2, 2015.
{{cite web}}: Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - ^ Staff (September 1, 2015). "Pentecopterus decorahensis: Ancient Giant Sea Scorpion Unearthed in Iowa". Sci-news.com. Retrieved September 2, 2015.
- ^ Lamsdell JC, Briggs DE, Liu HP, Witzke BJ, McKay RM (September 2015). "The oldest described eurypterid: a giant Middle Ordovician (Darriwilian) megalograptid from the Winneshiek Lagerstätte of Iowa". BMC Evolutionary Biology. 15: 169. doi:10.1186/s12862-015-0443-9. PMC 4556007. PMID 26324341.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Borenstein, Seth (31 August 2015). "Fossils show big bug ruled the seas 460 million years ago". AP News. Retrieved September 2, 2015.
{{cite news}}: Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - ^ Størmer L (1955). "Merostomata". Treatise on Invertebrate Paleontology, Part P Arthropoda 2, Chelicerata. p. 23.
- ^ a b c d e f g h i j "Palaeos Metazoa: Arthropoda: Chelicerata: Eurypterida: Eurypterida". 2007-08-13. Archived from the original on 2007-08-13. Retrieved 2017-12-17.
{{cite web}}: Unknown parameter|deadurl=ignored (|url-status=suggested) (help) - ^ a b Braddy SJ, Poschmann M, Tetlie OE (February 2008). "Giant claw reveals the largest ever arthropod". Biology Letters. 4 (1): 106–9. doi:10.1098/rsbl.2007.0491. PMC 2412931. PMID 18029297.
- ^ Shelton, Jim (August 31, 2015). "Meet Pentecopterus, a new predator from the prehistoric seas". Yale University. Retrieved September 1, 2015.
{{cite web}}: Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - ^ Tetlie, O. E. (2008). "Hallipterus excelsior, a Stylonurid (Chelicerata: Eurypterida) from the Late Devonian Catskill Delta Complex, and Its Phylogenetic Position in the Hardieopteridae". Bulletin of the Peabody Museum of Natural History. 49: 19–99. doi:10.3374/0079-032X(2008)49[19:HEASCE]2.0.CO;2.
- ^ a b c O'Connell, Marjorie. The Habitat of the Eurypterida.
{{cite book}}: Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - ^ Ortega-Hernández, J.; Legg, D. A.; Braddy, S. J. (2013). "The phylogeny of aglaspidid arthropods and the internal relationships within Artiopoda". Cladistics. 29: 15–45. doi:10.1111/j.1096-0031.2012.00413.x.
{{cite journal}}: Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - ^ Owen, Richard (1852). "Description of the impressions and footprints of the Protichnites from the Potsdam Sandstone of Canada". Quarterly Journal of the Geological Society of London. 8: 214–225. doi:10.1144/GSL.JGS.1852.008.01-02.26.
{{cite journal}}: Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - ^ Burling, Lancaster D. (1917). "Protichnites and Climactichnites. A critical study of some Cambrian trails". American Journal of Science. 44 (263): 387–398. doi:10.2475/ajs.s4-44.263.387.
{{cite journal}}: Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - ^ a b c d e f Lamsdell JC, Briggs DE, Liu HP, Witzke BJ, McKay RM (September 2015). "The oldest described eurypterid: a giant Middle Ordovician (Darriwilian) megalograptid from the Winneshiek Lagerstätte of Iowa". BMC Evolutionary Biology. 15: 169. doi:10.1186/s12862-015-0443-9. PMID 26324341.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ O'Connell, Marjorie. The Habitat of the Eurypterida.
{{cite book}}: Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - ^ Roy, Peter Van; Briggs, Derek E. G.; Gaines, Robert R. (2015-01-01). "The Fezouata fossils of Morocco; an extraordinary record of marine life in the Early Ordovician". Journal of the Geological Society: 2015–017. doi:10.1144/jgs2015-017. ISSN 0016-7649.
- ^ "Meet Pentecopterus, a new predator from the prehistoric seas". YaleNews. 2015-08-31. Retrieved 2018-01-05.
- ^ a b c d e f g h Dunlop JA, Penney D, Jekel D (2015). "A summary list of fossil spiders and their relatives." (PDF). World Spider Catalog. Natural History Museum Bern.
- ^ Mass Extinctions and Their Aftermath. Oxford University Press. 1997. p. 70. ISBN 978-0198549161.
- ^ Lamsdell JC, Braddy SJ (April 2010). "Cope's Rule and Romer's theory: patterns of diversity and gigantism in eurypterids and Palaeozoic vertebrates". Biology Letters. 6 (2): 265–9. doi:10.1098/rsbl.2009.0700. PMC 2865068. PMID 19828493.
- ^ a b Lamsdell, James C.; Braddy, Simon J.; Tetlie, O. Erik (2010-03-15). "The systematics and phylogeny of the Stylonurina (Arthropoda: Chelicerata: Eurypterida)". Journal of Systematic Palaeontology. 8 (1): 49–61. doi:10.1080/14772011003603564. ISSN 1477-2019.
{{cite journal}}: Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - ^ Benton MJ (2005). When life nearly died: the greatest mass extinction of all time. London: Thames & Hudson. ISBN 0-500-28573-X.
- ^ Tetlie, O. Erik (2007). "Distribution and dispersal history of Eurypterida (Chelicerata)" (PDF). Palaeogeography, Palaeoclimatology, Palaeoecology. 252 (3–4): 557–574. doi:10.1016/j.palaeo.2007.05.011. Archived from the original (PDF) on 2011-07-18.
{{cite journal}}: Unknown parameter|deadurl=ignored (|url-status=suggested) (help); Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - ^ Whyte MA (December 2005). "Palaeoecology: a gigantic fossil arthropod trackway". Nature. 438 (7068): 576. doi:10.1038/438576a. PMID 16319874.
{{cite journal}}: Cite has empty unknown parameter:|1=(help) - ^ a b Selden PA (1999). "Autecology of Silurian Eurypterids" (PDF). Special Papers in Palaeontology. 32. Palaeontological Association: 39–54. ISSN 0038-6804. Retrieved May 23, 2011.[permanent dead link]
- ^ Merck, John W. "The biomechanics of swimming". Department of Geology, University of Maryland. Retrieved May 23, 2011.
{{cite web}}: Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - ^ Merck, John W. "Eurypterids, arachnids, and the arthropod invasion of the land". Department of Geology, University of Maryland. Retrieved May 23, 2011.
{{cite web}}: Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - ^ Lamsdell, James C.; Braddy, Simon J.; Tetlie, O. E. (2009-09-01). "Redescription of Drepanopterus abonensis (Chelicerata: Eurypterida: Stylonurina) from the late Devonian of Portishead, UK". Palaeontology. 52 (5): 1113–1139. doi:10.1111/j.1475-4983.2009.00902.x. ISSN 1475-4983.
{{cite journal}}: Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - ^ a b c d Clarke, John Mason; Ruedemann, Rudolf. The Eurypterida of New York.
{{cite book}}: Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - ^ a b Garwood RJ, Dunlop J (2014). "Three-dimensional reconstruction and the phylogeny of extinct chelicerate orders". PeerJ. 2: e641. doi:10.7717/peerj.641. PMC 4232842. PMID 25405073.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Weygoldt P, Paulus HF (1979). "Untersuchungen zur Morphologie, Taxonomie und Phylogenie der Chelicerata". Zeitschrift für zoologische Systematik und Evolutionsforschung. 17 (2): 85–116, 177–200. doi:10.1111/j.1439-0469.1979.tb00694.x.
- ^ Versluys J, Demoll R (1920). "Die Verwandtschaft der Merostomata mit den Arachnida und den anderen Abteilungen der Arthropoda". Koninklijke Akademie van Wetenschappen Amsterdam. 23: 739–765.
- ^ Tetlie, O. Erik; Braddy, Simon J. (2004). "The first Silurian chasmataspid, Loganamaraspis dunlopi gen. et sp. nov. (Chelicerata: Chasmataspidida) from Lesmahagow, Scotland, and its implications for eurypterid phylogeny". Transactions of the Royal Society of Edinburgh: Earth Sciences. 94 (3): 227–234. doi:10.1017/S0263593300000638.
{{cite journal}}: Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - ^ Shultz, Jeffrey W. (2007). "A phylogenetic analysis of the arachnid orders based on morphological characters". Zoological Journal of the Linnean Society. 150 (2): 221–265. CiteSeerX 10.1.1.119.850. doi:10.1111/j.1096-3642.2007.00284.x.
{{cite journal}}: Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - ^ a b c Tetlie, O. Erik (2007). "Distribution and dispersal history of Eurypterida (Chelicerata)" (PDF). Palaeogeography, Palaeoclimatology, Palaeoecology. 252 (3–4): 557–574. doi:10.1016/j.palaeo.2007.05.011. Archived from the original (PDF) on 2011-07-18.
{{cite journal}}: Unknown parameter|deadurl=ignored (|url-status=suggested) (help); Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - ^ Romer AS (August 1933). "EURYPTERID INFLUENCE ON VERTEBRATE HISTORY". Science. 78 (2015): 114–7. doi:10.1126/science.78.2015.114. PMID 17749819.
- ^ a b Tetlie, O. Erik; Briggs, Derek E. G. (2009). "The origin of pterygotid eurypterids (Chelicerata: Eurypterida)". Palaeontology. 52 (5): 1141–1148. doi:10.1111/j.1475-4983.2009.00907.x.
{{cite journal}}: Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - ^ Briggs, D. E. G.; Fortey, R. A.; Clarkson, E. N. K. (1988). "Extinction and the fossil record of Arthropods". In G.P. Larwood (ed.). Extinction and survival in the fossil record. Oxford [Oxfordshire]: Published for the Systematics Association by Clarendon Press. pp. 171–209. ISBN 0198577087.
{{cite book}}: Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - ^ Ridley, Mark (2004). Evolution. Wiley-Blackwell. ISBN 978-1-4051-0345-9.
{{cite book}}: Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help)
Further reading
- Braddy, Simon J. (2001). "Eurypterid Palaeoecology: palaeobiological, ichnological and comparative evidence for a 'mass-moult-mate' hypothesis". Palaeogeography, Palaeoclimatology, Palaeoecology. 172 (1–2): 115–132. doi:10.1016/S0031-0182(01)00274-7.
{{cite journal}}: Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - Ciurca, Samuel J. (1998). "The Silurian Eurypterid Fauna". Retrieved July 25, 2004.
{{cite web}}: Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - Clarke, John M.; Rudolf, R. (1912). The Eurypterida of New York. Albany, New York: New York State Education Department.
{{cite book}}: Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - Gupta, Neal S.; Tetlie, O. Erik; Briggs, Derek E. G.; Pancost, Richard D. (2007). "The fossilization of eurypterids: a result of molecular transformation" (PDF). Palaios. 22 (4): 439–447. doi:10.2110/palo.2006.p06-057r.
{{cite journal}}: Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - Manning, Phillip L.; Dunlop, Jason A. (1995). "The respiratory organs of eurypterids" (PDF). Palaeontology. 38 (2): 287–297. Archived from the original (PDF) on 2012-03-09.
{{cite journal}}: Unknown parameter|deadurl=ignored (|url-status=suggested) (help); Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - Tetlie, O. Erik (2007). "Distribution and dispersal history of Eurypterida (Chelicerata)" (PDF). Palaeogeography, Palaeoclimatology, Palaeoecology. 252 (3–4): 557–574. doi:10.1016/j.palaeo.2007.05.011. Archived from the original (PDF) on 2011-07-18.
{{cite journal}}: Unknown parameter|deadurl=ignored (|url-status=suggested) (help); Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help) - Tetlie, O. Erik; Cuggy, Michael B. (2007). "Phylogeny of the basal swimming eurypterids (Chelicerata; Eurypterida; Eurypterina)". Journal of Systematic Palaeontology. 5 (3): 345–356. doi:10.1017/S1477201907002131.
{{cite journal}}: Unknown parameter|name-list-format=ignored (|name-list-style=suggested) (help)
External links
| ||
| Taxonomy |  | |
| Geochronology | ||
| Geography | ||
| Notable genera | ||
| Ichnogenera | ||
| Related groups | ||
| Related articles | ||